NERVOUS SYSTEM
The nervous system can be separated into parts based on structure and on function:
- structurally, it can be divided into the central nervous system (CNS) and the peripheral nervous system (PNS) (Fig. 1.33);
- functionally, it can be divided into somatic and visceral parts.
|
| The CNS is composed of the brain and spinal cord, both of which develop from the neural tube in the embryo.
|
| The
PNS is composed of all nervous structures outside the CNS that connect
the CNS to the body. Elements of this system develop from neural crest
cells and as outgrowths of the CNS. The PNS consists of the spinal and
cranial nerves, visceral nerves and plexuses, and the enteric system.
The detailed anatomy of a typical spinal nerve is described in Chapter 2, as is the way spinal nerves are numbered. Cranial nerves are described in Chapter 8. The details of nerve plexuses are described in chapters dealing with the specific regions in which the plexuses are located.
|
| The
parts of the brain are the cerebral hemispheres, the cerebellum, and
the brainstem. The cerebral hemispheres consist of an outer portion, or
the gray matter, containing cell bodies, an inner portion, or the white matter, made up of axons forming tracts or pathways, and the ventricles, which are spaces filled with cerebrospinal fluid (CSF).
|
| The
cerebellum has two lateral lobes and a midline portion. The components
of the brainstem are classically defined as the diencephalon, midbrain,
pons, and medulla. However, in common usage today, the term "brainstem"
usually refers to the midbrain, pons, and medulla.
|
| A further discussion of the brain can be found in Chapter 8.
|
The
spinal cord is the part of the CNS in the superior two- thirds of the
vertebral canal. It is roughly cylindrical in shape, and is circular to
oval in cross-section with a central canal. A further discussion of the
spinal cord can be found in Chapter 2.
|
Figure 1.34 Arrangement of meninges in the cranial cavity.
The meninges (Fig. 1.34)
are three connective tissue coverings that surround, protect, and
suspend the brain and spinal cord within the cranial cavity and
vertebral canal, respectively:
- the dura mater is the thickest and most external of the coverings;
- the arachnoid mater is against the internal surface of the dura mater;
- the pia mater is adherent to the brain and spinal cord.
|
| Between the arachnoid and pia mater is the subarachnoid space, which contains CSF.
|
| A further discussion of the cranial meninges can be found in Chapter 8 and of the spinal meninges in Chapter 2.
|
| Functional subdivisions of the CNS
|
Functionally, the nervous system can be divided into somatic and visceral parts.
- The somatic part (soma, from the Greek for
"body") innervates structures (skin and most skeletal muscle) derived
from somites in the embryo, and is mainly involved with receiving and
responding to information from the external environment.
- The visceral part (viscera,
from the Greek for "guts") innervates organ systems in the body and
other visceral elements, such as smooth muscle and glands, in peripheral
regions of the body. It is concerned mainly with detecting and
responding to information from the internal environment.
|
| Somatic part of the nervous system
|
The somatic part of the nervous system consists of:
- nerves that carry conscious sensations from peripheral regions back to the CNS; and
- nerves that innervate voluntary muscles.
|
Somatic nerves arise segmentally along the developing CNS in association with
somites, which are themselves arranged segmentally along each side of the neural tube (Fig. 1.35). Part of each somite (the
dermatomyotome)
gives rise to skeletal muscle and the dermis of the skin. As cells of
the dermatomyotome differentiate, they migrate into posterior (dorsal)
and anterior (ventral) areas of the developing body:
- cells that migrate anteriorly give rise to muscles of the limbs and trunk (hypaxial muscles) and to the associated dermis;
- cells that migrate posteriorly give rise to the intrinsic muscles of the back (epaxial muscles) and the associated dermis.
| Figure 1.35 Differentiation of somites in a "tubular" embryo. |
| Developing
nerve cells within anterior regions of the neural tube extend processes
peripherally into posterior and anterior regions of the differentiating
dermatomyotome of each somite (Fig. 1.36).
|
Simultaneously,
derivatives of neural crest cells (cells derived from neural folds
during formation of the neural tube) differentiate into neurons on each
side of the neural tube and extend processes both medially and laterally
(Fig. 1.37):
- medial processes pass into the posterior aspect of the neural tube;
- lateral processes pass into the differentiating regions of the adjacent dermatomyotome.
|
| Neurons that develop from neurons within the spinal cord are motor neurons and those that develop from neural crest cells are sensory neurons.
|
| Somatic
sensory and somatic motor fibers that are organized segmentally along
the neural tube become parts of all spinal nerves and some cranial
nerves.
|
| The clusters of sensory nerve cell bodies derived from neural crest cells and located outside the CNS form sensory ganglia. | | |
|

Figure 1.36 Somatic motor neurons.
Figure 1.37 Somatic sensory neurons. Blue lines indicate motor nerves and red lines indicate sensory nerves.
Generally, all sensory information passes into the posterior aspect of the spinal cord, and all motor fibers leave anteriorly.
| Somatic sensory neurons carry information from the periphery into the CNS and are also called somatic sensory afferents or general somatic afferents (GSAs).
The modalities carried by these nerves include temperature, pain,
touch, and proprioception. Proprioception is the sense of determining
the position and movement of the musculoskeletal system detected by
special receptors in muscles and tendons.
|
| Somatic motor fibers carry information away from the CNS to skeletal muscles and are also called somatic motor efferents or general somatic efferents (GSEs).
Like somatic sensory fibers that come from the periphery, somatic motor
fibers can be very long. They extend from cell bodies in the spinal
cord to the muscle cells they innervate.
|
Because
cells from a specific somite develop into the dermis of the skin in a
precise location, somatic sensory fibers originally associated with that
somite enter the posterior region of the spinal cord at a specific
level and become part of one specific spinal nerve (Fig. 1.38). Each spinal nerve therefore carries somatic sensory information from a specific area of skin on the surface of the body. A
dermatome is that area of skin supplied by a single spinal cord level, or on one side, by a single spinal nerve.

There
is overlap in the distribution of dermatomes, but usually a specific
region within each dermatome can be identified as an area supplied by a
single spinal cord
level. Testing touch in these autonomous zones in a conscious patient
can be used to localize lesions to a specific spinal nerve or to a
specific level in the spinal cord.
| Somatic
motor nerves that were originally associated with a specific somite
emerge from the anterior region of the spinal cord and, together with
sensory nerves from the same level, become part of one spinal nerve.
Therefore each spinal nerve carries somatic motor fibers to muscles that
originally developed from the related somite. A myotome is that portion of a skeletal muscle innervated by a single spinal cord level or, on one side, by a single spinal nerve.
|
Myotomes
are generally more difficult to test than dermatomes, because each
skeletal muscle in the body is usually innervated by nerves derived from
more than one spinal cord level (Fig. 1.39).

Visceral part of the nervous system
The visceral part of the nervous system, as in the somatic part, consists of motor and sensory components:
- sensory nerves monitor changes in the viscera;
- motor nerves mainly innervate smooth muscle, cardiac muscle, and glands.
|
| The visceral motor component is commonly referred to as the autonomic division of the PNS and is subdivided into sympathetic and parasympathetic parts.
|
| Like the somatic part of the nervous system, the visceral part is segmentally arranged and develops in a parallel fashion (Fig. 1.41).
|
Visceral sensory neurons
that arise from neural crest cells send processes medially into the
adjacent neural tube and laterally into regions associated with the
developing body. These sensory neurons and their processes, referred to
as
general visceral afferent fibers (
GVAs), are associated primarily with chemoreception, mechanoreception, and stretch reception.
Figure 1.40 Dermatomes (anterior view).
Visceral part of the nervous system
The visceral part of the nervous system, as in the somatic part, consists of motor and sensory components:
- sensory nerves monitor changes in the viscera;
- motor nerves mainly innervate smooth muscle, cardiac muscle, and glands.
|
| The visceral motor component is commonly referred to as the autonomic division of the PNS and is subdivided into sympathetic and parasympathetic parts.
|
| Like the somatic part of the nervous system, the visceral part is segmentally arranged and develops in a parallel fashion (Fig. 1.41).
|
Visceral sensory neurons
that arise from neural crest cells send processes medially into the
adjacent neural tube and laterally into regions associated with the
developing body. These sensory neurons and their processes, referred to
as
general visceral afferent fibers (
GVAs), are associated primarily with chemoreception, mechanoreception, and stretch reception.
Figure 1.41 Development of the visceral part of the nervous system.
Visceral
motor neurons that arise from cells in lateral regions of the neural
tube send processes out of the anterior aspect of the tube. Unlike in
the somatic part, these processes, containing general visceral efferent
fibers (GVEs), synapse with other cells, usually other visceral motor
neurons, that develop outside the CNS from neural crest cells that
migrate away from their original positions close to the developing
neural tube.
| The visceral motor neurons located in the spinal cord are referred to as preganglionic motor neurons and their axons are called preganglionic fibers; the visceral motor neurons located outside the CNS are referred to as postganglionic motor neurons and their axons are called postganglionic fibers.
|
| The cell bodies of the visceral motor neurons outside the CNS often associate with each other in a discrete mass called a ganglion.
|
| Visceral sensory and motor fibers enter and leave the CNS with their somatic equivalents (Fig. 1.42).
Visceral sensory fibers enter the spinal cord together with somatic
sensory fibers through posterior roots of spinal nerves. Preganglionic
fibers of visceral motor neurons exit the spinal cord in the anterior
roots of spinal nerves along with fibers from somatic motor neurons.
|
| Postganglionic
fibers traveling to visceral elements in the periphery are found in the
posterior and anterior rami (branches) of spinal nerves.
|
Visceral
motor and sensory fibers that travel to and from viscera form named
visceral branches that are separate from the somatic branches. These
nerves generally form plexuses from which arise branches to the viscera.

Figure 1.42 Basic anatomy of a thoracic spinal nerve.
Visceral motor and sensory fibers do not enter and leave the CNS at all levels (Fig. 1.43):
- in the cranial region, visceral components are associated with four of the twelve cranial nerves (CN III, VII, IX, and X);
- in the spinal cord, visceral components are associated mainly with spinal cord levels T1 to L2 and S2 to S4.
Visceral motor components associated with spinal levels T1 to L2 are termed
sympathetic. Those visceral motor components in cranial and sacral regions, on either side of the sympathetic region, are termed
parasympathetic:
- the sympathetic system innervates structures in peripheral regions of the body and viscera;
- the parasympathetic system is more restricted to innervation of the viscera only.
Figure 1.43 Parts of the CNS associated with visceral motor components.
Figure 1.44 Sympathetic part of the autonomic division of the PNS.
The
sympathetic part of the autonomic division of the PNS leaves
thoracolumbar regions of the spinal cord with the somatic components of
spinal nerves T1 to L2 (Fig. 1.44).
On each side, a paravertebral sympathetic trunk extends from the base
of the skull to the inferior end of the vertebral column
where the two trunks converge anteriorly to the coccyx at the ganglion
impar. Each trunk is attached to the anterior rami of spinal nerves and
becomes the route by which sympathetics are distributed to the periphery
and all viscera.
| Visceral
motor preganglionic fibers leave the T1 to L2 part of the spinal cord
in anterior roots. The fibers then enter the spinal nerves, pass through
the anterior rami and
into the sympathetic trunks. One trunk is located on each side of the
vertebral column (paravertebral) and positioned anterior to the anterior
rami. Along the trunk is a series of segmentally arranged ganglia
formed from collections of postganglionic neuronal cell bodies where the
preganglionic neurons synapse with postganglionic neurons. Anterior
rami of T1 to L2 are connected to the sympathetic trunk or to a
ganglion, by a white ramus communicans, which carries preganglionic sympathetic fibers and appears white because the fibers it contains are myelinated.
|
| Preganglionic
sympathetic fibers that enter a paravertebral ganglion or the
sympathetic trunk through a white ramus communicans may provide the
following.
|
| Peripheral sympathetic innervation at the level of origin of the preganglionic fiber
|
| Preganglionic
sympathetic fibers may synapse with postganglionic motor neurons in
ganglia associated with the sympathetic trunk, after which
postganglionic fibers enter the same anterior ramus and are distributed
with peripheral branches of the posterior and anterior rami of that
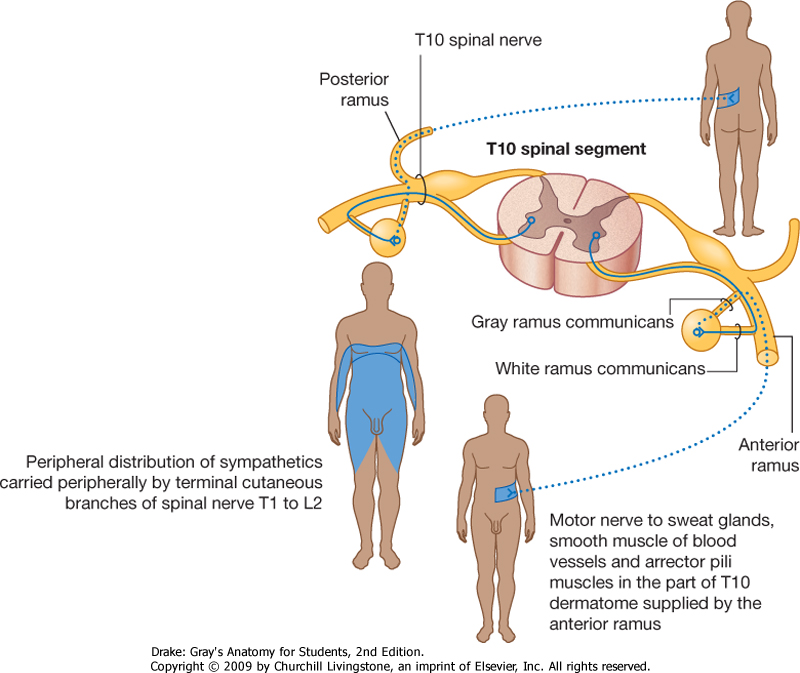
spinal nerve (Fig. 1.45). The fibers innervate structures at the periphery of the body in regions supplied by the spinal nerve. The gray ramus communicans
connects the sympathetic trunk or a ganglion to the anterior ramus and
contains the postganglionic sympathetic fibers. It appears gray because
postganglionic fibers are nonmyelinated. The gray ramus communicans is
positioned medial to the white ramus communicans.
|
| Peripheral sympathetic innervation above or below the level of origin of the preganglionic fiber
|
Preganglionic
sympathetic fibers may ascend or descend to other vertebral levels
where they synapse in ganglia associated with spinal nerves that may or
may not have visceral motor input directly from the spinal cord (i.e.,
those nerves other than T1 to L2) (Fig. 1.46
|
| The
postganglionic fibers leave the distant ganglia via gray rami
communicantes and are distributed along the posterior and anterior rami
of the spinal nerves. | | | | | | | | |

Figure 1.45 Course of sympathetic fibers that
travel to the periphery in the same spinal nerves in which they travel
out of the spinal cord.

Figure 1.46 Course of sympathetic nerves that
travel to the periphery in spinal nerves that are not the ones through
which they left the spinal cord.
The ascending and descending fibers, together with all the ganglia, form the paravertebral sympathetic trunk,
which extends the entire length of the vertebral column. The formation
of this trunk, on each side, enables visceral motor fibers of the
sympathetic part of the autonomic division of the PNS, which ultimately
emerge from only a small region of the spinal cord (T1 to L2), to be
distributed to peripheral regions innervated by all spinal nerves.
| White
rami communicantes only occur in association with spinal nerves T1 to
L2, whereas gray rami communicantes are associated with all spinal
nerves.
|
| Fibers
from spinal cord levels T1 to T5 pass predominantly superiorly, whereas
fibers from T5 to L2 pass inferiorly. All sympathetics passing into the
head have preganglionic fibers that emerge from spinal cord level T1
and ascend in the sympathetic trunks to the highest ganglion in the neck
(the superior cervical ganglion), where they synapse.
Postganglionic fibers then travel along blood vessels to target tissues
in the head, including blood vessels, sweat glands, small smooth muscles
associated with the upper eyelids, and the dilator of the pupil.
|
| Sympathetic innervation of thoracic and cervical viscera
|
Preganglionic
sympathetic fibers may synapse with postganglionic motor neurons in
ganglia and then leave the ganglia medially to innervate thoracic or
cervical viscera (
Fig. 1.47).
They may ascend in the trunk before synapsing, and after synapsing the
postganglionic fibers may combine with those from other levels to form
named visceral nerves, such as cardiac nerves. Often, these nerves join
branches from the parasympathetic system to form plexuses on or near the
surface of the target organ, for example, the cardiac and pulmonary
plexuses. Branches of the plexus innervate the organ. Spinal cord levels
T1 to T5 mainly innervate cranial, cervical, and thoracic viscera.
Sympathetic innervation of the abdomen and pelvic regions and the adrenals
| Preganglionic
sympathetic fibers may pass through the sympathetic trunk and
paravertebral ganglia without synapsing and, together with similar
fibers from other levels, form splanchnic nerves (greater, lesser, least, lumbar, and sacral), which pass into the abdomen and pelvic regions (Fig. 1.48). The preganglionic fibers in these nerves are derived from spinal cord levels T5 to L2.
|
| The
splanchnic nerves generally connect with sympathetic ganglia around the
roots of major arteries that branch from the abdominal aorta. These
ganglia are part of a large prevertebral plexus that also has input from
the parasympathetic part of the autonomic division of the PNS.
Postganglionic sympathetic fibers are distributed in extensions of this
plexus, predominantly along arteries, to viscera in the abdomen and
pelvis.
|
| Some
of the preganglionic fibers in the prevertebral plexus do not synapse
in the sympathetic ganglia of the plexus, but pass through the system to
the adrenal gland where they synapse directly with cells of the adrenal
medulla. These cells are homologues of sympathetic postganglionic
neurons and secrete adrenaline and noradrenaline into the vascular
system. | |


Parasympathetic system
The parasympathetic part of the autonomic division of the PNS (Fig. 1.49) leaves cranial and sacral regions of the CNS in association with:
- cranial nerves III, VII, IX, and X: III, VII, and IX carry
parasympathetic fibers to structures within the head and neck only,
whereas X (the vagus nerve) also innervates thoracic and most abdominal
viscera; and
- spinal nerves S2 to S4: sacral parasympathetic
fibers innervate inferior abdominal viscera, pelvic viscera, and the
arteries associated with erectile tissues of the perineum.
|
| page 48 |
 |
| page 49 |
| Like
the visceral motor nerves of the sympathetic part, the visceral motor
nerves of the parasympathetic part generally have two neurons in the
pathway. The preganglionic neurons are in the CNS, and fibers leave in
the cranial nerves.
|
| Sacral preganglionic parasympathetic fibers
|
| In the sacral region, the preganglionic parasympathetic fibers form special visceral nerves (the pelvic splanchnic nerves),
which originate from the anterior rami of S2 to S4 and enter pelvic
extensions of the large prevertebral plexus formed around the abdominal
aorta. These fibers are distributed to pelvic and abdominal viscera
mainly along blood vessels. The postganglionic motor neurons are in the
walls of the viscera. In organs of the gastrointestinal system,
preganglionic fibers do not have a postganglionic parasympathetic motor
neuron in the pathway; instead, preganglionic fibers synapse directly on
neurons in the ganglia of the enteric system.
|
| Cranial nerve preganglionic parasympathetic fibers
|
| The
preganglionic parasympathetic motor fibers in CN III, VII, and IX
separate from the nerves and connect with one of four distinct ganglia,
which house postganglionic motor neurons. These four ganglia are near
major branches of CN V. Postganglionic fibers leave the ganglia, join
the branches of CN V, and are carried to target tissues (salivary,
mucous, and lacrimal glands; constrictor muscle of the pupil; and
ciliary muscle in the eye) with these branches.
|
| The
vagus nerve [X] gives rise to visceral branches along its course. These
branches contribute to plexuses associated with thoracic viscera or to
the large prevertebral plexus in the abdomen and pelvis. Many of these
plexuses also contain sympathetic fibers.
|
| When present, postganglionic parasympathetic neurons are in the walls of the target viscera.
|
| Visceral sensory innervation (visceral afferents)
|
| Visceral sensory fibers generally accompany visceral motor fibers.
|
| Visceral sensory fibers accompany sympathetic fibers
|
| Visceral
sensory fibers follow the course of sympathetic fibers entering the
spinal cord at similar spinal cord levels. However, visceral sensory
fibers may also enter the spinal cord at levels other than those
associated with motor output. For example, visceral sensory fibers from
the heart may enter at levels higher than spinal cord level T1. Visceral
sensory fibers that accompany sympathetic fibers are mainly concerned
with detecting pain.
|
| Visceral sensory fibers accompany parasympathetic fibers
|
| Visceral sensory fibers accompanying parasympathetic fibers are carried mainly in IX and X and in spinal nerves S2 to S4.
|
| Visceral
sensory fibers in IX carry information from chemoreceptors and
baroreceptors associated with the walls of major arteries in the neck,
and from receptors in the pharynx.
|
| Visceral sensory fibers in X include those from cervical viscera, and major vessels and viscera in the thorax and abdomen.
|
| Visceral sensory fibers from pelvic viscera and the distal parts of the colon are carried in S2 to S4.
|
| Visceral
sensory fibers associated with parasympathetic fibers primarily relay
information to the CNS about the status of normal physiological
processes and reflex activities.
|
The
enteric nervous system consists of motor and sensory neurons and their
support cells, which form two interconnected plexuses, the myenteric and submucous nerve plexuses, within the walls of the gastrointestinal tract (Fig. 1.50). Each of these plexuses is formed by:
- ganglia, which house the nerve cell bodies and associated cells; and
- bundles of nerve fibers, which pass between ganglia and from the ganglia into surrounding tissues.
|
| Neurons
in the enteric system are derived from neural crest cells originally
associated with occipitocervical and sacral regions. Interestingly, more
neurons are reported to be in the enteric system than in the spinal
cord itself.
|
| Sensory
and motor neurons within the enteric system control reflex activity
within and between parts of the gastrointestinal system. These reflexes
regulate peristalsis, secretomotor activity, and vascular tone. These
activities can occur independently of the brain and spinal cord, but can
also be modified by input from preganglionic parasympathetic and
postganglionic sympathetic fibers.
|
Sensory information from the enteric system is carried back to the CNS by visceral sensory fibers.
Nerve plexuses
| Nerve
plexuses are either somatic or visceral and combine fibers from
different sources or levels to form new nerves with specific targets or
destinations (Fig. 1.51). Plexuses of the enteric system also generate reflex activity independent of the CNS.
|
Major
somatic plexuses formed from the anterior rami of spinal nerves are the
cervical (C1 to C4), brachial (C5 to T1), lumbar (L1 to L4), sacral (L4
to S4), and coccygeal (S5 to Co) plexuses. Except for spinal nerve T1,
the anterior rami of thoracic spinal nerves remain independent and do
not participate in plexuses.
Visceral plexuses
| Visceral
nerve plexuses are formed in association with viscera and generally
contain efferent (sympathetic and parasympathetic) and afferent
components (Fig. 1.51).
These plexuses include cardiac and pulmonary plexuses in the thorax,
and a large prevertebral plexus in the abdomen anterior to the aorta,
which extends inferiorly onto the lateral walls of the pelvis. The
massive prevertebral plexus supplies input to and receives output from
all abdominal and pelvic viscera.
|
| Referred pain
occurs when sensory information comes to the spinal cord from one
location, but is interpreted by the CNS as coming from another location
innervated by the same spinal cord level. Usually, this happens when the
pain information comes from a region, such as the gut, which has a low
amount of sensory output. These afferents converge on neurons at the
same spinal cord level that receive information from the skin, which is
an area with a high amount of sensory output. As a result, pain from the
normally low output region is interpreted as coming from the normally
high output region. |
| Pain is most often
referred from a region innervated by the visceral part of the nervous
system to a region innervated, at the same spinal cord level, by the
somatic side of the nervous system. |
Pain can also be
referred from one somatic region to another. For example, irritation of
the peritoneum on the inferior surface of the diaphragm, which is
innervated by the phrenic nerve, can be referred to the skin on the top
of the shoulder, which is innervated by other somatic nerves arising at
the same spinal cord level.





 Nerve plexuses
Nerve plexuses
 Visceral plexuses
Visceral plexuses

{kind=link}
{kind=link}




{kind=link}
No comments:
Post a Comment